1. Introduction
Mahogany, Swietenia macrophylla King (Meliaceae) is one of the most valuable large, deciduous Neotropical tree species on the world market (Gullison et al., 1996; Degen et al., 2012; Sebbenn et al., 2012). Its wood has excellent aesthetic, physical, and mechanical characteristics, remarkable workability, and is used in the manufacture of luxury furniture, interior decoration for civil construction, and musical instruments (Carvalho, 2006; André et al., 2008). The geographic range of S. macrophylla extends from southern Mexico to the southern Amazon basin in Bolivia, Brazil, and Peru (Lamb, 1966). In adulthood, individuals can reach a diameter at breast height > 2 m and a tree height of up to 50 m (Gullison et al., 1996), but population density is generally low, <1 tree/ha (Grogan et al., 2008). Logging of S. macrophylla began in the 16th century in Central American populations (Mejía et al., 2008), but large-scale harvesting began in South American populations in the 20th Century (Grogan et al., 2008), where due to over-exploitation during the 1970s and 1980s resulted in a decline in their natural populations (Alcalá et al., 2015). In Peru, the species has been exploited since the 1920s (Mejía et al., 2008). As a result, many natural S. macrophylla populations have become spatially isolated, resulting in a loss of genetic diversity (Novick et al., 2003; Breed et al., 2012; Degen et al., 2012; Sebbenn et al., 2012; Alcalá et al., 2015; Oliveira et al., 2020; Andrade et al., 2022). Due to that, S. macrophylla was included on the list of endangered species of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) Appendix II (CITES, 2019).
Across the geographic range of S. macrophylla natural distribution, demand for its timber has exceeded available supply due to extinction or decreases in natural population sizes throughout its range (Reynel et al., 2003; Campos et al., 2009; Castro et al., 2019; Andrade et al., 2022; Arlacon-Méndez et al., 2023). The future of mahogany as a commercial timber species is at risk due to over-exploitation and deforestation (Verwer et al., 2008). For this reason, the use of the species in commercial reforestation and degraded area restoration programs has been gaining prominence in various places across the Americas (Barrena & Vargas, 2004; Kurihara et al., 2005). However, insects such as Hypsipyla grandella Zeller (Lepidoptera: Pyralidae) have been identified as the main pest and the most important barrier to establishing and developing S. macrophylla commercial plantations (Grijpma & Gara, 1976; Lunz et al., 2009). The caterpillar attacks the terminal bud of the plant causing bifurcations (Grijpma & Gara, 1976; Lunz et al., 2009), or it can feed on the fruits and seeds of the plant (Castro et al., 2019).
Hypsipyla grandella is difficult to control, which is why multiple methods were tested with different results, even though they could significantly reduce populations, although even low population levels could cause important damage to the plantation, among the most commonly used ones are silvicultural management, chemical, bioecological control, integrated systems, resistance to hosting and combinations between methods, are some of them (Álvarez et al., 2021; Guerra-Arevalo et al., 2022; Dueñas, 2022; Borges et al., 2022; Ávila-Arias et al., 2023; Domínguez et al., 2023; García et al., 2024; Ávila-Arias et al., 2025).
One way to address this problem is to establish commercial plantations with seedlings that are resistant to H. grandella attacks (Newton et al., 1993; Mayhew & Newton, 1998; Schabel et al., 1999). To develop a selection strategy, the three aspects of coexistence, prevention, and sustainability must be considered, alongside the use of resistant or tolerant trees based on the antixenosis of H. grandella to the Meliaceae family throughout the Americas (Jiménez, 2007). In terms of improving tree response to H. grandella attacks, the terminal bud of the plant can be eliminated by pruning so that it does not bifurcate or emit several regrowth sprouts, with subsequent selection of the most vigorous plants (Navarro et al., 2004; Revilla-Chávez et al., 2024).
In this context, the use of techniques such as the predictive decapitation of S. macrophylla regrowth can help genetic selection by identifying trees with greater tolerance to H. grandella attacks, resulting in improved commercial plantation viability (Cornelius, 2009). Based on this technique, studies have observed genetic differences at the individual level, where S. macrophylla from Panama produced an average of 2.2 regrowth sprouts per tree after decapitation with scissors, while those from Costa Rica had 3.1 regrowth sprouts per tree (Hilje & Cornelius, 2001). In the case of Meliaceae species such as Cedrela sp. and S. macrophylla, the number of regrowth sprouts produced after an H. grandella attack represents an important component of attack tolerance, as species show a wide range of responses, from a single regrowth to 10 or more (Hilje & Cornelius, 2001). Thus, simulated attacks using scissors can be conducted across different genotypes early in the nursery phase for later establishment in the field. The results suggest that it is feasible to identify genotypes that are more tolerant to attack by using the decapitation test (Hilje & Cornelius, 2001; Revilla-Chávez et al., 2024).
The aim of this study was to assess the vegetative regrowth of S. macrophylla plants from four different families to simulated attacks by H. grandella in a nursery and examine the possibility of early selection of individuals more suitable for the development of forest plantations in the Peruvian Amazon. We emphasize that the present study represents an exploratory investigation (pilot study) to assess the possibility of early selection of S. macrophylla individuals for vegetative regrowth before H. grandella attack in the nursery phase. This is not an empirical study aimed at using the selected plants in a program of genetic improvement of the species, as a progeny test with only four S. macrophylla families is insufficient to conduct selection between and within progenies in a breeding program.
2. Methodology
The study was carried out in the Forest Nursery at the Faculty of Forestry and Environmental Sciences of the National University of Ucayali, Manantay district, Coronel Portillo Province, Ucayali Region, Peru. For the study, open-pollinated seeds from four seed-trees were sampled at four locations: Juanjui (UTM 326823E, 9055066N; elevation of 283 m) and Sisa (326878E; 8767893N; elevation of 430 m), representing families from the San Martin region; and Primavera (558797E; 8779586N; elevation of 164 m) and Unión (559748E; 8801935N; elevation of 164 m), representing families from the Ucayali region (Figure 1). On September 6, 2018, 175 seeds per family were sown directly into polyethylene bags with fertile substrate, composed of two volumes of black soil, one of sand and one of worm humus, with a texture of 40% silt, 30% clay and 30% sand. Of the sown seeds, 27 Juanjui seeds, 16 Sisa seeds, 143 Primavera seeds, and 25 Unión seeds germinated. On December 29, 2018, when the plants reached an average height of 30 cm, 50% of the crowns of 77 plants selected for their best phenotypic characteristics in the nursery were decapitated with pruning shears to simulate an attack by H. grandella (Figure 2). A completely randomized design with unbalanced repetitions was applied, with 22 from the Juanjui family, 22 from the Primavera family, 22 from the Unión family, and 11 from Sisa. After decapitation, the plants were evaluated at 194 days of age. Plant height (H, cm), plant base height (HB, cm), stem diameter (D, cm), number of regrowth sprouts (NR), sum of regrowth length (RL, cm), regrowth basal area (BA, mm2), number of branches (NB), and number of leaves (NL), were evaluated until robust regrowth sprouts were obtained.

In relation to the provenances, Sisa projected toward the positive side of Dim1, suggesting a greater influence of traits associated with a high number of regrowth (NR). Juanjui was located toward the negative side of Dim1, indicating contrasting traits, likely related to greater plant height (H) and base height (HB). Primavera was positioned near the origin of the coordinate plane, indicating intermediate phenotypic values for most variables. Unión, meanwhile, separated along Dim2, suggesting particular traits possibly related to stem diameter (D) and other secondary variables. Overall, PCA facilitated the identification of key traits that differentiate the provenances, with NR, RL, and NL contributing the most to the observed variation. These results provide valuable information for selecting materials with greater regrowth potential and for understanding the genetic structure among the provenances studied.
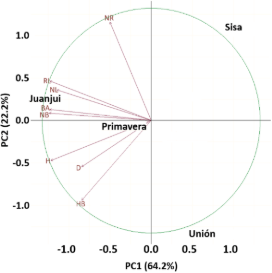
Figure 4. Principal component analysis (PCA) for the Juanjui, Primavera, Unión and Sisa provenances for the variables plant height (H, cm), plant base height (HB, cm), stem diameter (D, cm), number of regrowth sprouts (NR), sum of regrowth length (RL, cm), regrowth basal area (BA, mm2), number of branches (NB), and number of leaves (NL).
The results of the Principal Component Analysis (PCA) show clear patterns of phenotypic differentiation among the evaluated provenances, based on variables primarily related to regrowth behavior and plant structural growth. Sisa's separation toward positive values of the first principal component (Dim1) suggests greater regrowth vigor, characterized by a greater number of regrowth (NR) and greater total regrowth length (RL), which could be associated with adaptive strategies in the face of environmental disturbances. In contrast, Juanjui's position on the negative side of Dim1 indicates a phenotypic profile centered on larger structural size, such as plant height (H) and base height (HB), but with a lower propensity for regrowth. This pattern could reflect adaptations to more stable environments, where height growth represents a competitive advantage.
Primavera's proximity to the origin suggests a provenance with intermediate behavior, while Unión, with its separation along Dim2, could represent a particular phenotypic variability, possibly associated with stem diameter (D) or traits related to bole robustness. These findings are consistent with recent studies highlighting the importance of phenotypic variability in genetic conservation strate-gies and the selection of superior materials (Zhou et al., 2021; Silva et al., 2023). The identification of key traits, such as the number of shoots and the sum of shoot lengths, provides a solid basis for future breeding programs aimed at enhancing the resilience and productivity of the study populations. Furthermore, the combined use of genetic and phenotypic correlations, along with PCA, proves to be an effective tool for exploring intraspecific diversity and for outlining management and conservation strategies tailored to the genetic characteristics of each provenance.
4. Conclusions
The conclusions should be interpreted with caution, as the study was based on a limited sample of four open-pollinated families of Swietenia macrophylla. After simulating an attack by Hypsipyla grandella, genetic differences were observed in stem diameter, regrowth length, and basal area, suggesting potential for genetic improvement through early selection. Regrowth length emerged as the most relevant trait, showing genetic correlations with plant height and diameter, enabling indirect selection at the nursery stage. Furthermore, height, diameter, and basal area of regrowth showed moderate genotypic determination coefficients, supporting their use in mass selection. Principal Component Analysis (PCA) revealed structured differentiation among provenances, with plant height, basal height, number of regrowth, and regrowth length contributing most to multivariate discrimination, indicating underlying genetic patterns that could be strategically exploited in breeding and conservation programs.
Acknowledgments
We thank the National University of Ucayali for financing this project, the Peruvian Amazon Research Institute for technical support, and Dr. Jonathan Cornelius for the scientific suggestions in the present study. We also thank Dr. Evelyn R. Nimmo for assistance with English language editing.
Contribution from authors
J. M. Revilla-Chávez: Writing – original draft. R. D. Manturano-Pérez: Acquisition of financing, Methodology and review. A. C. Gonzales-Alvarado: Data curation. R. Casas-Reátegui: Conceptualization, formulation or evolution of general research goals and objectives. L. I. Briceño Dueñas: Data curation. J. A. Mori-Vásquez: Methodology, Development or design of methodology; Model Creation. A. M. Sebbenn: Proofreading and editing, Formal analysis.
ORCID
J. M. Revilla-Chávez https://orcid.org/0000-0001-5978-5146
R. D. Manturano-Pérez https://orcid.org/0000-0002-5063-8593
A. C. Gonzales-Alvarado https://orcid.org/0000-0003-3793-0271
R. Casas-Reátegui https://orcid.org/0000-0003-3781-166X
L. I. Briceño Dueñas https://orcid.org/0009-0003-1857-2517
J. A. Mori-Vásquez https://orcid.org/0000-0003-0570-8369
A. M. Sebbenn https://orcid.org/0000-0003-2352-0941
References
Abarquez, A., Bush, D., Ata, J., Tolentino, E.L. Jr., & Gilbero, D. (2015). Early growth and genetic variation of mahogany (Swietenia macrophylla) in progeny tests planted in Northern Mindanao, Philippines. Journal of Tropical Forest Science, 27, 314-324.
André, T., Lemes, M. R., Grogan, J., & Gribel, R. (2008). Post-logging loss of genetic diversity in a mahogany (Swietenia macrophylla King, Meliaceae) population in Brazilian Amazonia. Forest Ecology and Management, 255, 430-345. https://doi.org/10.1016/j.foreco.2007.09.055
Alcalá, R. E., De La Cruz, S., & Gutiérrez-Granados, G. (2015). Genetic structure and genetic diversity of Swietenia macrophylla in areas subjected to selective logging in Quintana Roo, Mexico. Botanical Sciences, 93, 819-828. https://doi.org/10.17129/botsci.256
Andrade, R. L., Pico-Mendozab, J., Morillo, E., Buitrón, J., Meneses, S., Navarrete, B., Pinoargote, M., & Carrasco, B. (2022). Molecular characterization of mahogany tree (Swietenia macrophylla King, Meliaceae) in the remnant natural forest of Ecuador. Neotropical Biodiversity, 18, 222-228. https://doi.org/10.1080/23766808.2022.2080334
Arlacon-Méndez, M., Maselli, S., van Zonneveld, M., Loo, J., Snook, L., Oliva, A., Franco, A., & Duminil, J. (2023). Implications of community forest management for the conservation of the genetic diversity of big-leaf mahogany (Swietenia macrophylla King, Meliaceae) in the Maya Biosphere Reserve, Petén, Guatemala. Trees, Forests and People, 11, 100362. https://doi.org/10.1016/j.tfp.2022.100362
Álvarez, D., Zuleta, D., Saldamando, C., & Lobo-Echeverri, T. (2021). Selective activity of Carapa guianensis and Swietenia macrophylla (Meliaceae) against the corn and rice strains of Spodoptera frugiperda (Lepidoptera, Noctuidae). International Journal of Pest Management, 70(3), 409–422. https://doi.org/10.1080/09670874.2021.1981484
Ávila-Arias, C., González-González, W., Chichilla-Mora, O., & Meza-Picado, V. (2023). Desempeño juvenil de clones de caoba (Swietenia macrophylla King) en Pococí de Limón, Costa Rica. Revista Forestal Mesoamericana Kurú, 20(47), 35-47. http://dx.doi.org/10.18845/rfmk.v20i47.6817
Ávila-Arias, C. E., Chinchilla Mora, O., Meza Picado, V., Fonseca González, W., & Arias Aguilar, D. (2025). Respuesta de clones de caoba (Swietenia macrophylla King) a la fertilización química en invernadero. Colombia Forestal, 28(1), e21987. https://doi.org/10.14483/2256201X.21987
Barrena, V., & Vargas, C. (2004). Informe de la Autoridad Científica CITES, La caoba en el Perú. Universidad Nacional Agraria La Molina - Facultad de Ciencias Forestales, Lima, 31p.
Breed, M. F., Gardner, M. G., Ottewell, K. M., Navarro, C. M., & Lowe, A. J. (2012). Shifts in reproductive assurance strategies and inbreeding costs associated with habitat fragmentation in Central American mahogany. Ecology Letters, 15, 444-452. https://doi.org/10.1111/j.1461-0248.2012.01752.x
Borges, R., Boff, M. I. C., Mantovani, A., Borges, M., Laumann, R. A., & Blassioli-Moraes, M. C. (2022). Why shading cedar (Cedrela fissilis) reduces damage caused by mahogany shoot borer, Hypsipyla grandella (Zeller)?. Forest Ecology and Management, 504, 119853. https://doi.org/10.1016/j.foreco.2021.119853
Castro, M. T., Montalvão, S. C. L., Peronti, A. L. B. G., & Monnerat, R. G. 2019. Ocorrência da cochonilha Phenacoccus solenopsis Tinsley (Hemiptera, Pseudococcidae) em mudas de mogno (Swietenia macrophylla King). Entomo Brasilis, 12, 41-43. https://doi.org/10.12741/ebrasilis.v12i1.791
Campos, L., Collazos, H., Reátegui, K., Valcárcel, H., & López, J. (2009). Evaluación económica de plantaciones de "caoba" Swietenia macrophylla en el departamento de San Martín. Instituto de Investigaciones de la Amazonía Peruana, Iquitos, Perú, 44p.
Carvalho, P. E. R. (2006). Espécies arbóreas brasileiras. Embrapa Informação Tecnológica, Brasília, Brazil. 629p.
CITES. (2019). Appendix II. https://www.cites.org/resources/species.html
Cornelius, J.P. (2009). The utility of the predictive decapitation test as a tool for early genetic selection for Hypsipyla tolerance in big-leaf mahogany (Swietenia macrophylla King). Forest Ecology and Management, 257, 1815-1821. https://doi.org/10.1016/j.foreco.2009.02.003
Crista, L., Radulov, I., Crista, F., Imbrea, F., Manea, D. N., Boldea, M., Gergen, I., Ienciu, A. A., & Lațo, A. (2024). Utilizing Principal Component Analysis to Assess the Effects of Complex Foliar Fertilizers Regarding Maize (Zea mays L.) Productivity. Agriculture, 14(8), 1428. https://doi.org/10.3390/agriculture14081428
Domínguez-Liévano, A., Aguilera Rodríguez, M., Espinosa Zaragoza, S., Aldrete, A., & Wong Villarreal, A. (2023). Sustratos y fertilización para producir planta de Swietenia macrophylla King y Tabebuia donnell-smithii Rose en charolas. Revista Mexicana de Ciencias Forestales, 14(77), 56-75. https://doi.org/10.29298/rmcf.v14i77.1332
Degen, B., Ward, S. E., Lemes, M. R., Navarro, C., Cavers, S., & Sebbenn, A. M. (2012). Verifying the geographic origin of mahogany (Swietenia macrophylla King) with DNA-fingerprints. Forensic Science International, Genetics, 7, 55-62. https://doi.org/10.1016/j.fsigen.2012.06.003
Dueñas-López, M. A. (2022). Hypsipyla grandella (mahogany shoot borer). CABI Compendium. https://doi.org/10.1079/cabicompendium.28356
García, C. C., Tovar, D. C., Morales, A. V., Bolaños, I. A., Llanderal, V. D. C., & de Abreu-Loureiro, G. A. H. (2024). The Effect of application methods and chemical insecticides for the attack control of Hypsipyla grandella Zeller. Revista de Ciências Agro-Ambientais, 22(1), 46-61. https://doi.org/10.30681/rcaa.v22i1.12009
Grijpma, P., & Gara, R. I. (1976). Studies on the shootborer Hypsipyla grandella (Zeller), II. Host preference of the larva. Turrialba, 20, 241–247. https://repositorio.iica.int/handle/11324/13999
Grogan, J., Jennings, S. B., Landis, R M., Schulze, M., Baima, A. M. V., Lopes, J. C. A., Norghauer, J. M., Oliveira, L. R., Pantoja, F., Pinto, D., Silva, J. N. M., Vidal, E., & Zimmerman, B. L. (2008). What loggers leave behind, Impacts on big-leaf mahogany (Swietenia macrophylla) commercial populations and potential for post-logging recovery in the Brazilin Amazon. Forest Ecology and Management, 255, 269-281. https://doi.org/10.1016/j.foreco.2007.09.048
Guerra-Arévalo, H., Celis-Espinoza. D., Díaz-Visitación, A. I., Vásquez-Vela, A. L. M., Arévalo-López, L. A., García-Soria, D. G., Revilla-Chávez, J. M., et al. (2022). Azadirachta indica y Jatropha curcas reducen el ataque de Hypsipyla grandella Zéller en Swietenia macrophylla King plantada en sistemas agroforestales. Scientia Forestalis, 50, e3674. https://doi.org/10.18671/scifor.v50.10
Gullison, R. E., Panfil, S. N., Strouse, J. J., & Hubbell, S. P. (1996). Ecology and management of mahogany (Swietenia macrophylla King.) in the Chimanes Forest, Beni, Bolivia. Botanical Journal of the Linnean Society, 122, 9-34. https://doi.org/10.1111/j.1095-8339.1996.tb02060.x
Hilje, L., & Cornelius, J. (2001). Es inmanejable Hypsipyla grandella como plaga forestal? CATIE. Manejo Integrado de Plagas, 61, i-iv. https://repositorio.catie.ac.cr/handle/11554/6958
Jiménez, Q. (2007). Monografía de la familia Meliaceae. In Hammel, B.E., Grayum, M.H., Herrera, C., & Zamora, N. (eds) Manual de las plantas de Costa Rica. Missouri Botanical Garden, Instituto Nacional de Biodiversidad, Museo Nacional de Costa Rica, p. 575-614.
Kurihara, D. L., Imaña-Encinas, J., & Paula, E. (2005). Levantamento da arborização do campus da Universidade de Brasilia. CERNE, 11, 127-136.
Lamb, F. B. (1966). Mahogany of Tropical America, Its ecology and management. University of Michigan Press, Ann Arbor, Michigan, 220p.
Lunz, A. M., Thomazini, M. J., Moraes, M. C. B., Neves, E. J. M., Batistca, T. F. C., Degenhardt, J., Sousa, L. A., & Ohashi, O. S. (2009). Hypsipyla grandella em Mogno (Swietenia macrophylla), Situação atual e perspectivas. Pesquisa Florestal Brasileira, 59, 45-55.
Mayhew, J. E., & Newton, A .C. (1998). The silviculture of mahogany. CABI Publications, Wallingford, 226p. https://doi.org/10.1023/A,1006233910888
Mejía, E., Buitrón, X., Peña-Claros, M., & Grogan, J. (2008). Big-leaf mahogany (Swietenia macrophylla) in Peru, Bolivia and Brazil. NDF Workshop Case Studies. WG1-Trees, Case Study 4 Swietenia macrophylla. Mexico, 36p. https://doi.org/10.1111/1365-2664.12210
Navarro, C., & Hernández, G. (2004). Progeny test analysis and population differentiation of Mesoamerican mahogany (Swietenia macrophylla). Agronomía Costarricense, 28, 37-51.
Navarro, C., Montagnini, F., & Hernández, G. (2004). Genetic variability of Cedrela odorata Linnaeus, Results of early performance of provenances and families from Mesoamerica grown in association with coffee. Forest Ecology and Management, 192, 217-227. https://doi.org/10.1016/j.foreco.2004.01.037
Novick, R. R., Lemes, M. R., Navarro, C., Caccone, A., & Bermingham, E. (2003). Genetic structure of Mesoamerican populations of big-leaf mahogany (Swietenia macrophylla) inferred from microsatellite analysis. Molecular Ecology, 12, 2885-2893. https://doi.org/10.1046/j.1365-294X.2003.01951.x
Newton, A., Baker, P., Ramnarine, S., Mesén, J. F., & Leakey, R. R. B. (1993). The mahogany shoot borer, Prospects for control. Forest Ecology and Management, 57, 301-328. https://doi.org/10.1016/0378-1127(93)90179-Q
Oliveira, S. S., Campos, T., Sebbenn, A. M., & d`Oliveira, M. V. V. (2020). Using spatial genetic structure of a population of Swietenia macrophylla King to integrate genetic diversity into management strategies in Southwestern Amazon. Forest Ecology and Management, 464, 118040. https://doi.org/10.1016/j.foreco.2020.118040
Pan, S., Liu, Z., Han, Y., Zhang, D., Zhao, X., Li, J., & Wang, K. (2024). Using the Pearson’s correlation coefficient as the sole metric to measure the accuracy of quantitative trait prediction, Is it sufficient? Frontiers in Plant Science, 15, 1480463. https://doi.org/10.3389/fpls.2024.1480463
Resende, M. D. V. (2016). Software Selegen – REML/BLUP, a useful tool for plant breeding. Crop Breeding and Applied Biotechnology, 16, 330-339. https://doi.org/10.1590/1984-70332016v16n4a49
Resende, M. D. V. (2002). Genética biométrica e estatística no melhoramento de plantas perenes. EMBRAPA Informação Tecnológica, Brasília, 975p.
Revilla-Chávez, J. M., de Moraes, M. A., Revilla-Macedo, J. J., Vergaray-Rengifo, W. F., Mego-Pérez, J. A., Saldaña-Dominguez, H. S., Vigo-Ampuero, E. S., Gonzales-Alvarado, A. C., Manturano-Pérez, R. D., Casas-Reátegui, R., Mori-Vasquez, J. A., & Sebbenn, A. M. (2024). Correlaciones, parámetros genéticos y fenotípicos en rasgos cuantitativos y cualitativos de Swietenia macrophylla en Ucayali, Perú. Scientia Agropecuaria, 15(3), 409-417. https://doi.org/10.17268/sci.agropecu.2024.030
Reynel, C., Pennington, R.T., Pennington, T. D., Flores, C., & Daza, A. (2003). Árboles útiles de la Amazonía peruana y sus usos, Tarea Gráfica Educativa, Perú, 509p.
Ríos-Saucedo, J. C., Acuña-Carmona, E., Cancino-Cancino, J., Rubilar-Pons, R., Corral-Rivas, J. J., & Rosales-Serna, R. (2017). Dinámica de brotación y densidad básica de la madera en rebrotes de tres especies dendroenergéticas. Agrociencia, 51, 215–227.
Schabel, H. G., Hilje, L., Nair, K. S. S., & Varma, R. V. (1999). Economic entomology in tropical forest plantations: An update. Journal of Tropical Forest Science, 11, 303–315. https://www.jstor.org/stable/43582530.
Sebbenn, A. M. (2002). Número de árvores matrizes e conceitos genéticos na coleta de sementes para reflorestamentos com espécies nativas. Revista do Instituto Florestal, 14, 115-132. https://doi.org/10.24278/2178-5031.2002142412
Sebbenn, A. M., Licona, J. C., Mostacedo, B., & Degen, B. (2012). Gene flow in an overexploited population of Swietenia macrophylla King (Meliaceae) in the Bolivian Amazon. Silvae Genetica, 61, 212-220. https://doi.org/10.1515/sg-2012-0027
Silva, T. L., Martins, E. P., & Ferreira, A. (2023). Genetic diversity and phenotypic plasticity in response to environmental gradients. Plant Ecology, 224(2), 151-165. https://doi.org/10.1007/s11258-023-01234-7
Verwer, C., Pena-Claros, M., Van-Der-Staak, D., Ohlson-Kiehn, K., & Sterck, F. (2008). Silviculture enhances the recovery of overexploited mahogany Swietenia macrophylla. Journal Applied Ecology, 45, 1770-1779. https://doi.org/10.1111/j.1365-2664.2008.01564.x
Wightman, K. E., Ward, S. E., Haggar, J. P., Santiago, B. R., & Cornelius, J. P. (2008). Performance and genetic variation of big-leaf mahogany (Swietenia macrophylla King) in provenance and progeny trials in the Yucatan Peninsula of Mexico. Forest Ecology and Management, 255, 346-355. https://doi.org/10.1016/j.foreco.2007.09.056
Zhou, H., Yang, L., Zhang, J., & Wang, X. (2021). Exploring genetic variability and phenotypic correlations in perennial forage species. Euphytica, 217(6), 1-15. https://doi.org/10.1007/s10681-021-02831-9