RESUMEN
Se determinó los principales aspectos tróficos de Ethmidium maculatum “machete” en la región La Libertad, durante el año 2016. Se analizó una muestra de 362 estómagos llenos. Se determinó la dinámica trófica a través del análisis de los índices de vacuidad (I.V.) y repleción (I.R). Para obtener la composición trófica se utilizó los métodos frecuencia de ocurrencia, numérico y volumétrico (biomasa húmeda de presas). La dinámica alimentaria indicó dos periodos tróficos importantes y la mayor llenura de estómagos fue en abril. La dieta estuvo conformada por 39 presas agrupadas en cinco ítems presa. Las diatomeas pelágicas fueron las más frecuentes y numerosas, sin embargo, en cuanto al porcentaje de biomasa húmeda, fue mayor en el ítem copépodo. Los resultados constituyen un aporte significativo al conocimiento de la dinámica alimentaria y alimento de E. maculatum, no obstante, se necesitan estudios a mayor escala espacial y temporal, que permitan un análisis a nivel de hábitat trófico de peces planctófagos.
Palabras claves: Ethmidium maculatum, dinámica trófica, alimento y hábitos alimentarios, región La Libertad.
ABSTRACT
The main trophic aspects of Ethmidium maculatum “machete” were determined in La Libertad Region, during 2016. A sample of 362 full stomachs was analyzed. The trophic dynamics was determined through the analysis of the vacuity (I.V.) and repletion (I.R) indices. To obtain the trophic composition, the frequency of occurrence, numerical and volumetric methods (wet biomass of preys) were used. The food dynamics indicate two important trophic periods, and the greatest stomach filled were on April. The diet consisted of 39 preys grouped into five items. Pelagic diatoms were the most frequent and numerous; however, in terms of the percentage of wet biomass, it was higher in the item copepods. The results constitute a significant contribution to the knowledge of the food and feeding dynamics of E. maculatum; however, studies on a larger spatial and temporal scale are needed, which allow an analysis at the trophic habitat level of planktophagous fish.
Keywords: Ehtmidium maculatum, trophic dynamics, food and feeding habits, La Libertad region
DOI: http://dx.doi.org/10.17268/rebiol.2020.40.02.09
1. INTRODUCCIÓN
La biología trófica y los hábitos alimenticios de los peces, permiten comprender el papel biológico que desempeñan en su hábitat, a través de las relaciones entre el alimento con el crecimiento, desove, ritmo circadiano y el desarrollo de su ciclo de vida (Nikolsky, 1963; Wootton, 1979, 1990; Link y Almeida, 2000; Khan’ko et al., 2009; Yuneva et al., 2016). Además, se pueden conocer interacciones ecológicas fundamentales en la estructuración de comunidades marinas (Choy et al., 2015; Shaw et al., 2016).
Ethmidium maculatum “machete” es un pez marino que pertenece a la familia Clupeidae y recibe otros nombres comunes como “machetillo” y “machuelo”. Su distribución geográfica abarca desde el Golfo de Guayaquil en Ecuador, por el norte, hasta Puerto de Talcahuano por el sur (0°-37°S) (Chirichigno, 1974; Whitehead, 1985). Esta especie es considerada un recurso marino de aceptación comercial en la pesca artesanal peruana, utilizada en la gastronomía debido a su alto valor nutricional en proteínas, sodio, potasio, calcio, magnesio y hierro (IMARPE, 1996).
En la región La Libertad, el machete es uno de los principales recursos pesqueros desembarcados (Atoche, 2016), por lo que este estudio aporta mayor información sobre el alimento y hábitos alimentarios de E. maculatum. Pocos son los estudios realizados para esta especie; se ha estudiado la Biología y Pesquería de machete (Atoche y Culquichicón 2013). En los aspectos tróficos, según Atoche (2014), el grado de digestibilidad semidigerido puede variar entre 80% y 94%; y el grupo zooplancton (Calanus sp) es el más representativo según los métodos de estimación porcentual y frecuencia de ocurrencia; sin embargo, estas estimaciones son cualitativas y a criterio del investigador, por lo que es necesario obtener indicadores cuantitativos en cuanto a ecología trófica de esta especie. Por ello, el objetivo de este estudio fue determinar los principales aspectos tróficos como la dinámica alimentaria, composición y abundancia de presas presentes en el contenido estomacal de machete en la región La Libertad durante el año 2016.
2. MATERIAL Y MÉTODOS
El área de estudio se localizó en la zona marino costera de la región La Libertad, donde se realiza la actividad pesquera artesanal, en los puntos de desembarque: Puerto Pacasmayo, Puerto Malabrigo, Caleta Huanchaco, Puerto Salaverry y Caleta Puerto Morín. Se analizó 14 muestras de febrero a diciembre del 2016; éstas procedieron de 14 zonas de pesca (Fig. 1). Estas capturas fueron realizadas por embarcaciones de madera denominadas “bote” de 2 a 4 t de capacidad de bodega, utilizando redes cortina de 76,2 mm de tamaño de malla.
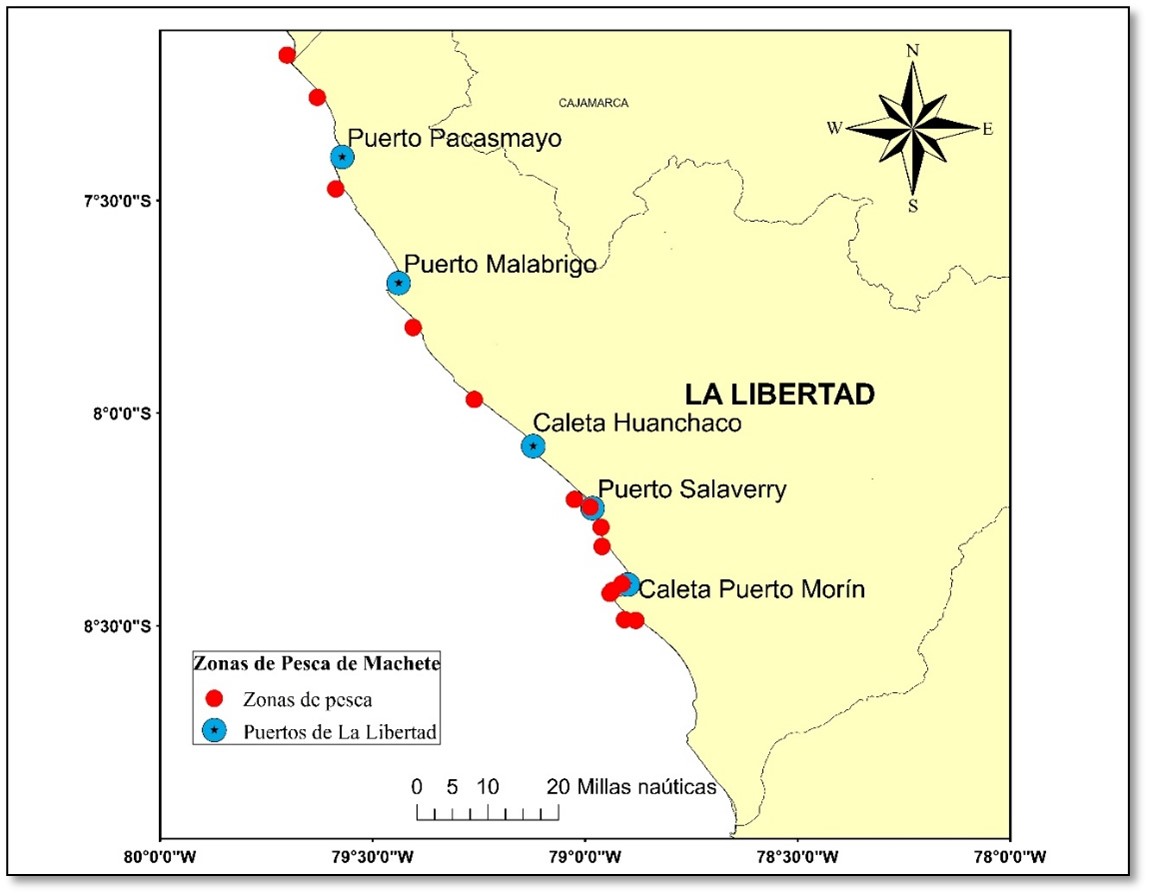
Figura 1. Ubicación geográfica de la zona de estudio.
Se analizó 481 ejemplares de E. maculatum de 22 a 32 cm de longitud total. Del total de ejemplares, 362 estómagos fueron llenos, de los cuales se obtuvieron datos de peso de estómago lleno (PELL), peso de estómago vacío (PEV) y peso de contenido estomacal (PCE) en gramos; para ello se utilizó una balanza marca KERN de 0,01 g de sensibilidad.
La dinámica alimentaria se calculó a través de los índices de vacuidad y de repleción o llenado. El índice de vacuidad permitió establecer los periodos de mayor o menor actividad alimentaria (Pitt, 1973; Ramos, 1979); y el índice de repleción, para conocer el comportamiento alimenticio, es decir, los periodos donde los estómagos tienen mayor contenido de alimento (De Silva, 1973; Hyslop, 1980).

Donde:
I.V.: Índice de vacuidad
EV: Número de estómagos vacíos
ET: Número de estómagos totales analizados
I.R.: Índice de repleción o llenado
Para determinar el alimento de E. maculatum se empleó los métodos de frecuencia de ocurrencia, con el que se determinó la presencia o ausencia de cada presa en el estómago, más información cuantitativa a través del método numérico (Tresierra y Culquichicón, 1995) y el volumétrico, procedimiento propuesto por el protocolo interno del Laboratorio de Ecología trófica del IMARPE (Ruíz, 2016).
Para el análisis del contenido estomacal se analizó el primer estómago por intervalo de talla. Los ejemplares fueron agrupados por intervalos de talla de 4 cm de amplitud, y para el análisis de las presas se unió el contenido de los cuatro estómagos (máximo) en una luna de reloj. Posterior a ello, se filtró la muestra con el uso de dos tamices, uno con tamaño de malla de 75μm para fitoplancton y de 300 μm para zooplancton.
Para reconocer los organismos presa se utilizó un microscopio binocular Zeiss - Primo Star, en el cual se analizó las muestras de los contenidos estomacales a un aumento de 10x; además de un estereoscopio Leica - 6sd; y bibliografía especializada, para fitoplancton (Cupp, 1943; Balech, 1988) y para zooplancton (Santander, 1981; Boltovskoy, 1981). Las presas fueron organizadas en ítems alimentarios o presa, para un mejor entendimiento de los patrones; en fitoplancton, las presas de clasificaron en: diatomeas bentónicas, diatomeas pelágicas, dinoflagelados y silicoflagelados; en zooplancton, solo se consideró un ítem alimentario, copépodos; miscelánea, fue referencial, mas no fue considerado como ítem presa, sino como agrupación de otras presas no importantes.
La componente fitoplanctónica, se analizó en un frasco de 20 ml, luego se llevó una alícuota (0,1 ml) a una lámina portaobjeto; cubriéndola con una laminilla; realizándose un conteo total del campo. La componente zooplanctónica; se diluyó en un vaso de precipitación, llevándolo a 100 ml y con ayuda de una pipeta se tomó una alícuota de 10 ml, la cual fue vertida en una cámara Bogorov, donde se realizó el conteo respectivo.
Para estimar la biomasa húmeda por presa según el método volumétrico, se utilizó factores de conversión de biovolúmenes individuales de fitoplancton y zooplancton (Sun y Liu 2003; Taylor, 2008) y se siguió el procedimiento propuesto por el protocolo interno del Laboratorio de Ecología trófica del IMARPE (Ruíz, 2016). Posterior a ello, se aplicó la prueba estadística de Kruskal Wallis para verificar si existe diferencia significativa en la biomasa húmeda de ítems alimentarios por mes y por rangos de longitud total (variación ontogénica).
3. RESULTADOS
Para reconocer los niveles de conocimiento y su relación con las prácticas de fotoprotección se establecieron rangos para el nivel de conocimiento y las prácticas de fotoprotección.
El índice de vacuidad indicó dos periodos tróficos. De marzo a junio hay menor actividad trófica con respecto al último semestre de ese año, donde los valores fueron inferiores a 16,5%. Los mayores valores se obtuvieron en parte del verano y otoño, en marzo (34,7%), en abril (71,4%) y junio (29,4%). Luego se observó una disminución hasta agosto y noviembre, donde se alcanzó un índice de vacuidad del 0%. Finalmente, en diciembre el índice de vacuidad fue de 9,38% y comienza a incrementar nuevamente, lo que hace indicar un ciclo trófico (Fig. 2).
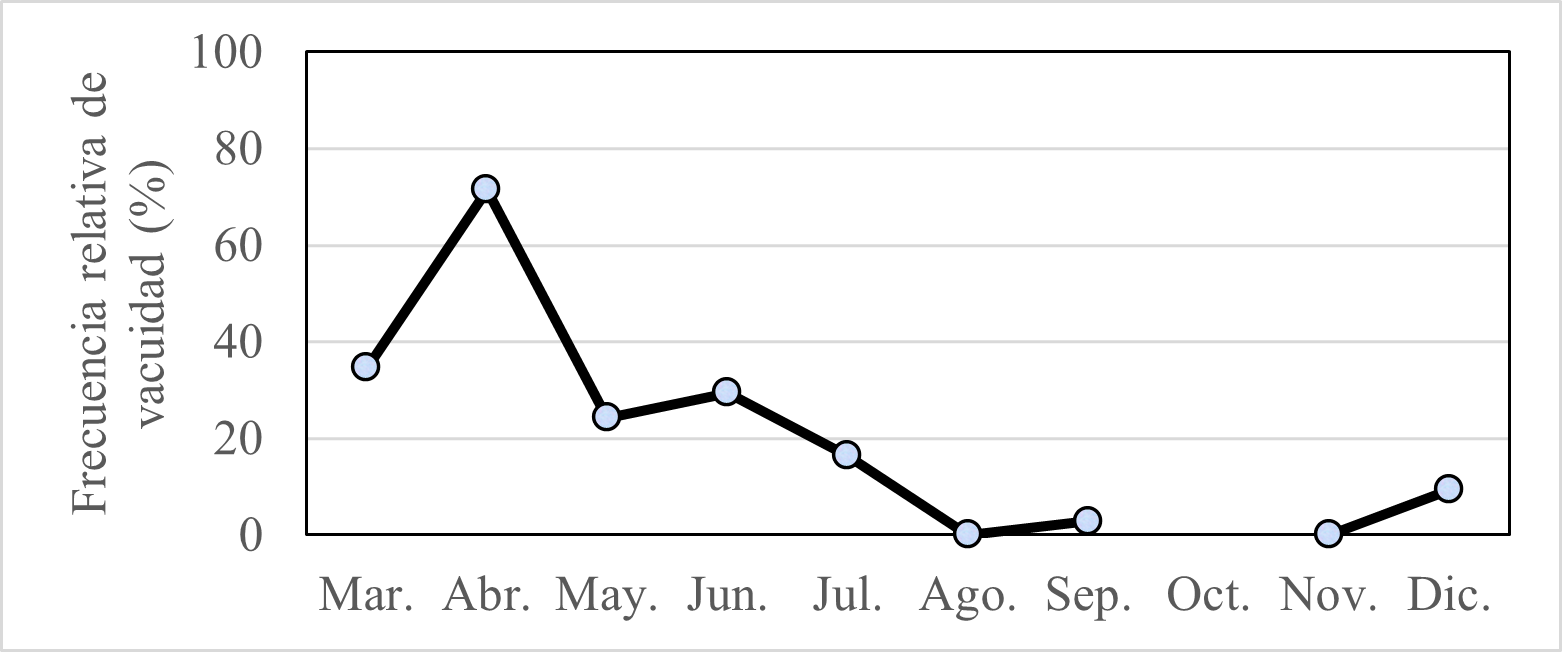
Figura 2. Índice de vacuidad de Ethmidium maculatum “machete”
El índice de repleción en marzo tuvo su mayor valor (0,7%). Posterior a ese mes, los valores no tuvieron mayor variación, salvo un segundo pico de 0,42%. El promedio anual fue de 0,4% (Figura 3)
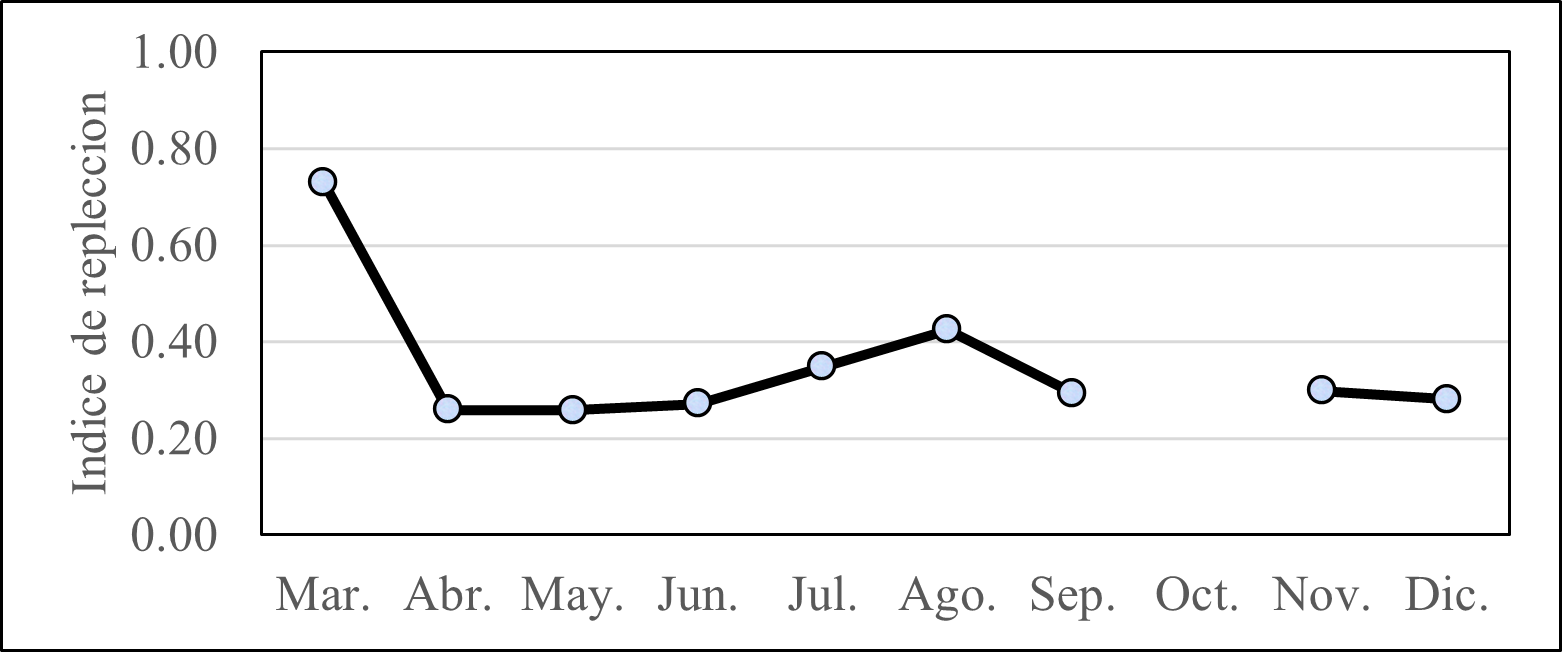
Figura 3. Índice de repleción de E. maculatum “machete”
La dieta de E. maculatum estuvo conformada por 39 presas. Estos ítems fueron constituidos de la siguiente manera: 3 géneros de diatomeas bentónicas, 9 géneros de diatomeas pelágicas, 4 géneros y 7 especies de dinoflagelados, 1 género de silicoflagelado, 8 géneros de copépodos, y 5 de miscelánea. En el ítem miscelánea se observó la presencia del Orden Euphausiacea, Clase Ostracoda, huevos de anchoveta, huevos de crustáceo y larvas de cirrípedos.
La dieta de E. maculatum según el método de frecuencia de ocurrencia indicó que los ítems alimentarios más frecuentes en todo el periodo de estudio fueron copépodos (33,6%) y diatomeas pelágicas (29,6%). Otros ítems alimentarios presentes fueron dinoflagelados, diatomeas bentónicas y silicoflagelados; y no se observó un patrón marcado en la variación de los ítems alimentarios (Fig. 4).
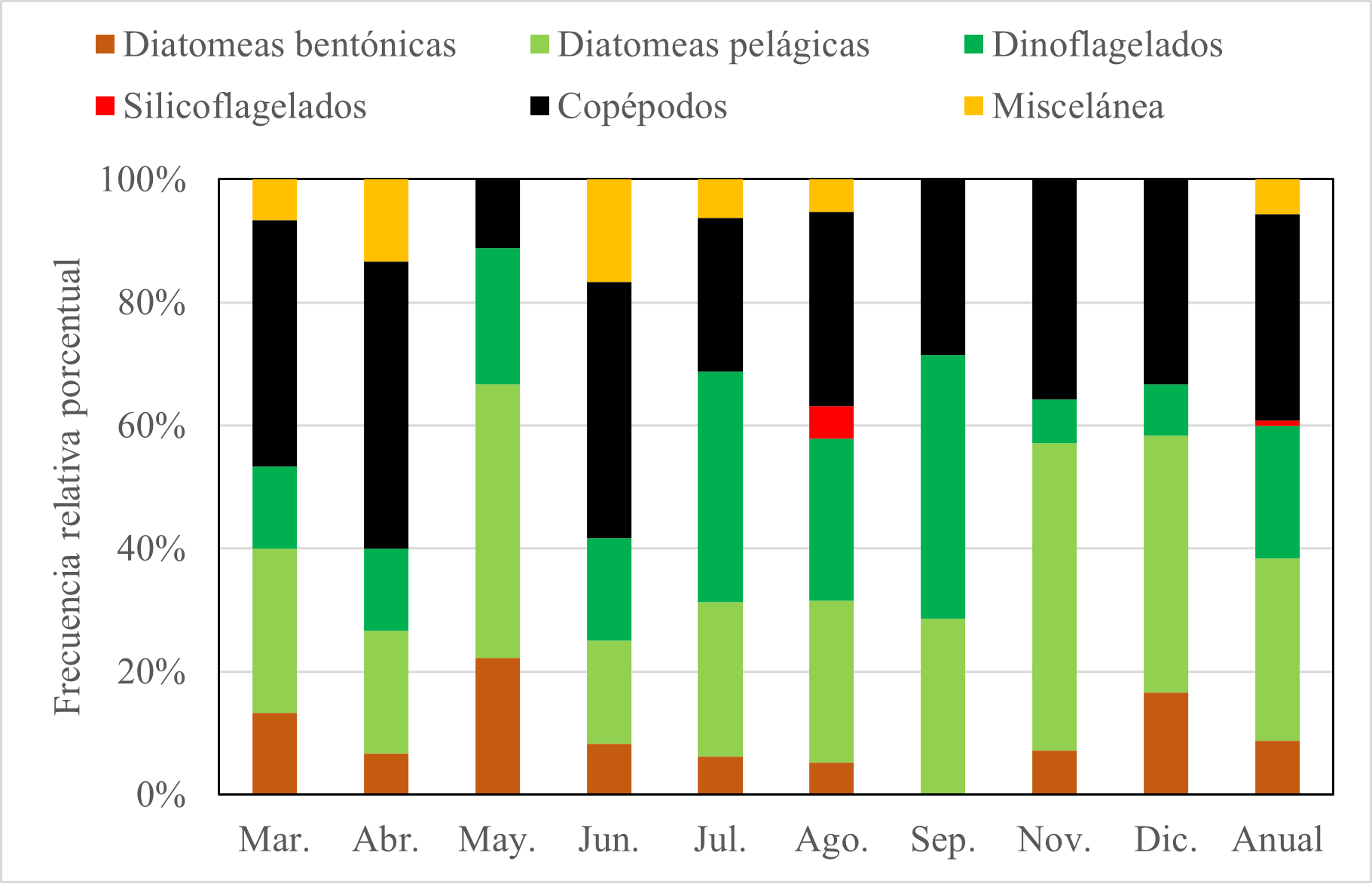
Figura 4. Variación mensual de ítems alimentarios de E. maculatum mediante el método de frecuencia de ocurrencia.
La dieta de E. maculatum según el método numérico indicaron que el ítem alimentario más abundante en todo el periodo de estudio fue las diatomeas pelágicas (55,5%); y hay patrones de variación para diatomeas pelágicas, copépodos y dinoflagelados. En el caso de las diatomeas pelágicas, se observó que los mayores valores correspondieron al periodo mayo-agosto. En el caso de los copépodos se observó que los mayores valores se dieron en marzo-abril y noviembre diciembre. Además, también se observó un patrón para dinoflagelados, donde los valores de este ítem no tuvieron mayor variación, salvo en septiembre, donde fue la presa más abundante (39,0%) (Fig. 5).
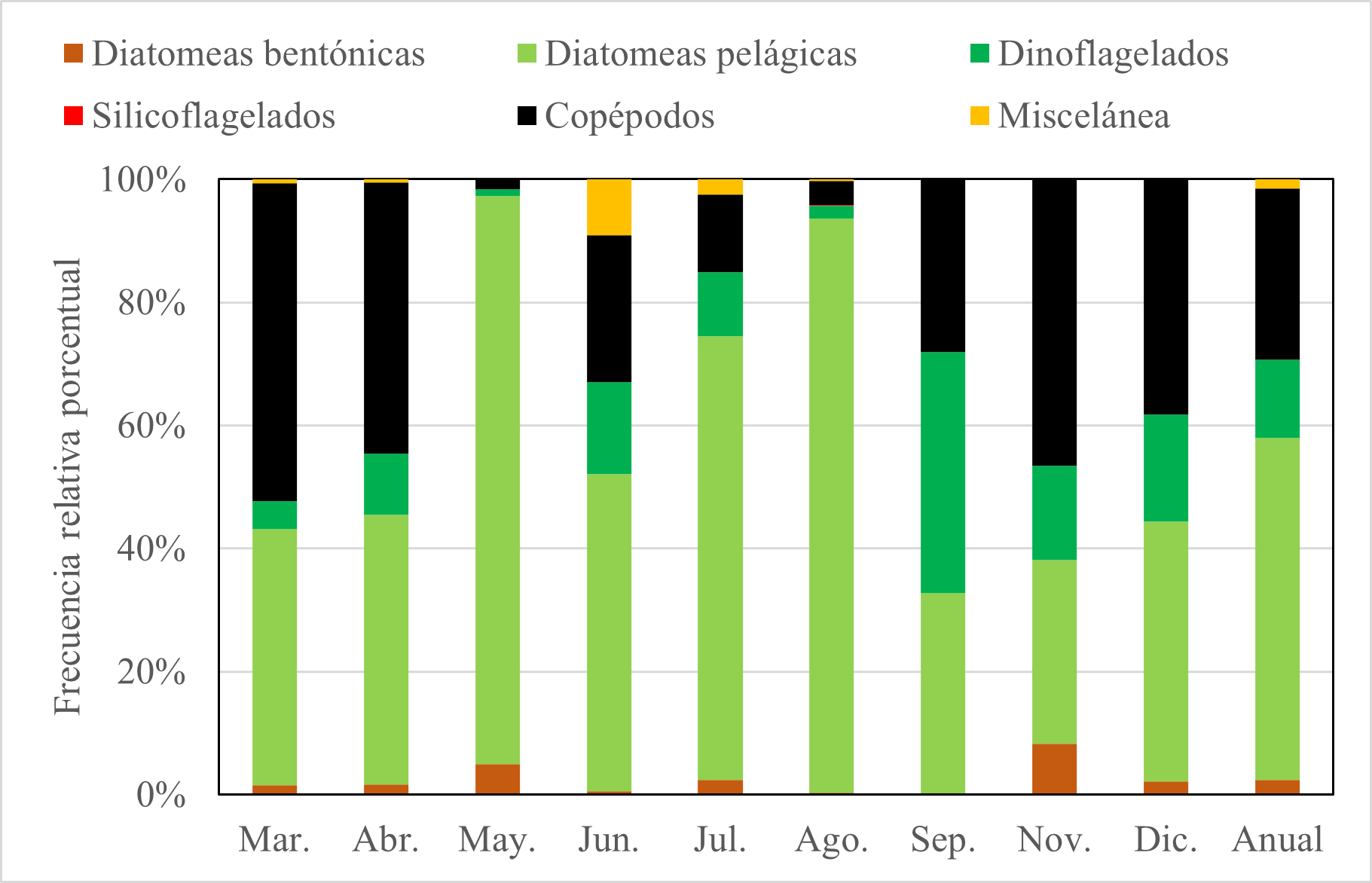
Figura 5. Variación mensual de ítems alimentarios de E. maculatum mediante el método numérico.
La dieta de E. maculatum según el método volumétrico indicó que el ítem alimentario más abundante en todo el periodo de estudio fue los copépodos (97,2%); y hay un patrón de variación para copépodos. Este patrón indicó que los copépodos predominaron en todo el año, pero en el periodo mayo-agosto se observó menores porcentajes (Fig. 6). Esto se verificó con un análisis no paramétrico, que indicó que no hay evidencia estadística para demostrar diferencias entre el consumo de diatomeas pelágicas (p valor: 0,525), diatomeas bentónicas (p valor: 0,244) y dinoflagelados (p valor: 0,990) en función al tiempo (mes); sin embargo, para el caso de los copépodos (p valor: 0,004) si hay diferencias, por lo que no es consumido en la misma cantidad en todos los meses.
Las presas con mayor importancia en porcentajes de biomasa húmeda en todo el año fueron: Oithona sp (49,4%), Calanus sp (20,6 %), Paracalanus sp (8,1%), Coscinodicus sp (5,1 %) y Oncaea sp (4,63%). Esto demuestra que la dieta de machete en función al método volumétrico, tuvo preferencia por el copépodo Oithona sp.
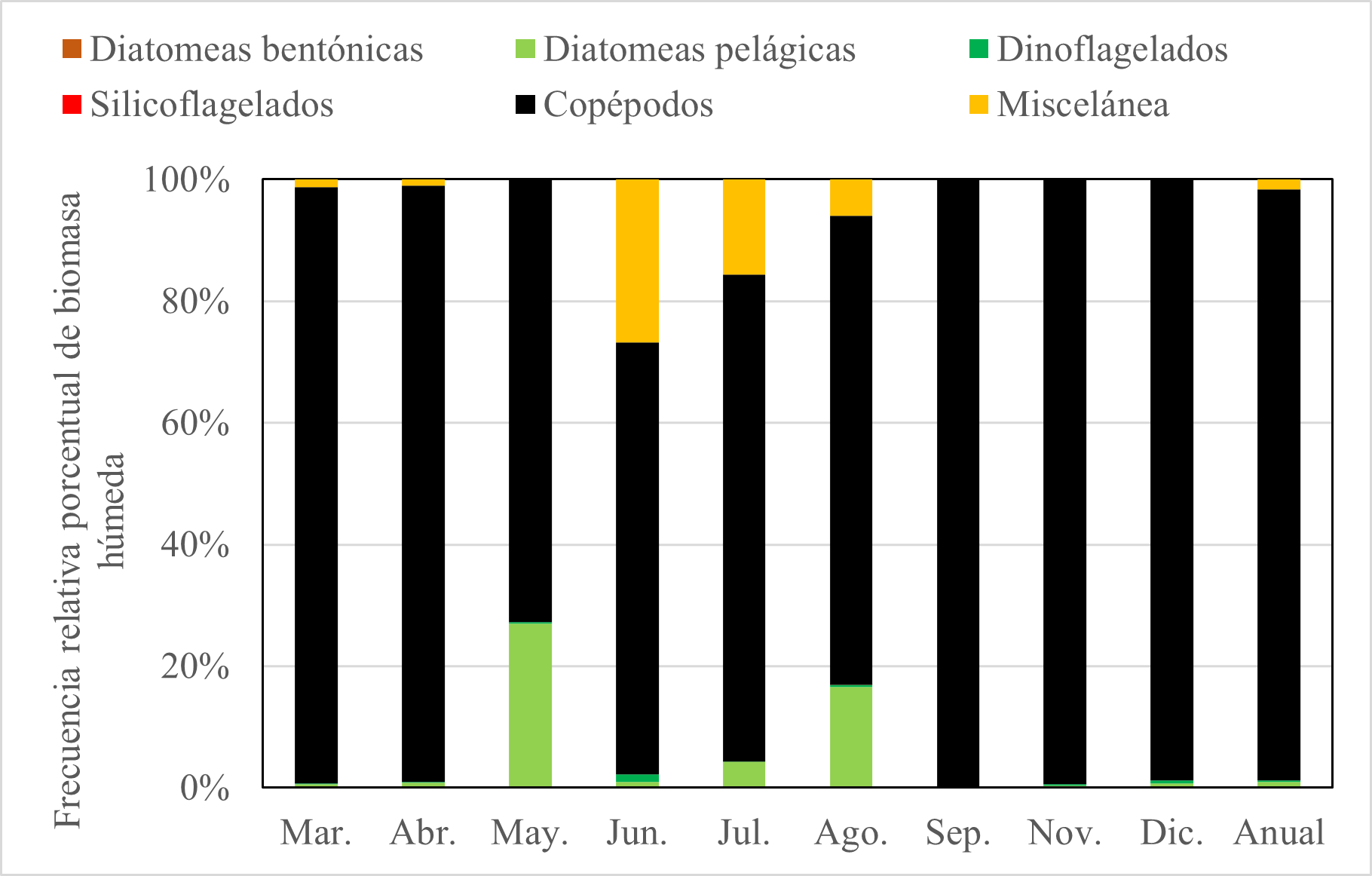
Figura 6. Variación mensual de ítems alimentarios de E. maculatum mediante el método volumétrico.
La dieta de E. maculatum según intervalos de longitud total (cm) no presentó un patrón definido y no hubo evidencia estadística para demostrar variación ontogénica en el consumo diatomeas pelágicas (p. valor: 0,997), diatomeas bentónicas (p. valor: 0,486), dinoflagelados (p. valor: 0,175) y copépodos (p. valor: 0,656). Los intervalos de longitud fueron conformados de 21 a 24 cm, de 25 a 28 cm y de 29 a 32cm de longitud total. Se observó que en el intervalo de talla 21- 24 cm, el 87,0% fueron copépodos y el 8,3% miscelánea; de 25-28 cm el porcentaje de copépodos fue 76,4% y diatomeas pelágicas fueron 12,0%; de 29-32cm el porcentaje de copépodos fue 94,5% y las diatomeas pelágicas 3,9% (Fig. 7).
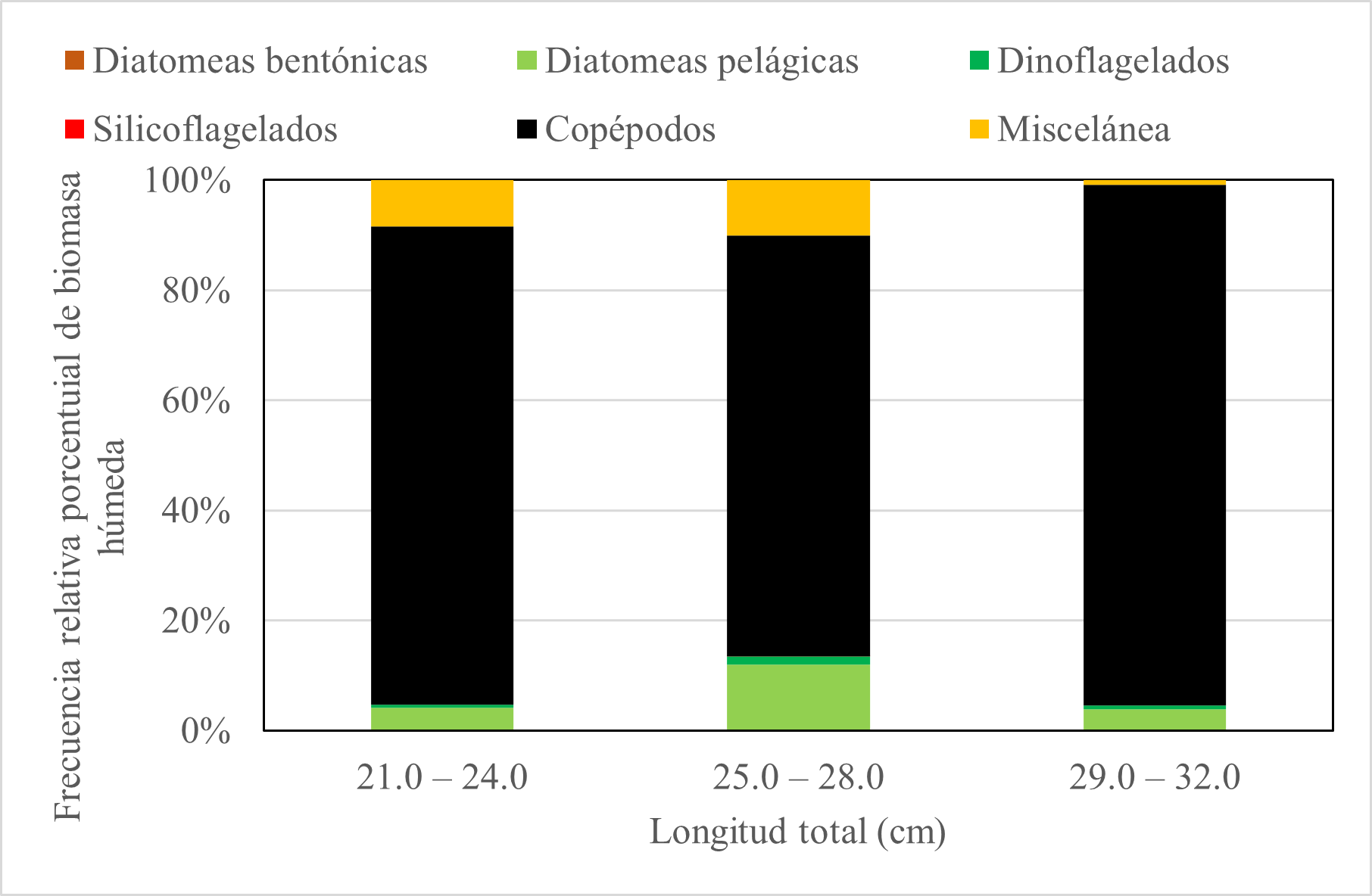
Figura 7. Variación ontogénica de la dieta de E. maculatum.
4. DISCUSIÓN
La dinámica alimentaria de machete a través del índice de vacuidad indicó que se relaciona con el periodo de mayor reproducción de la especie, que ocurre en invierno y primavera (Atoche y Culquichicón, 2013). A medida que la especie tiene mayor actividad reproductiva, se alimenta menos.
El promedio del índice de repleción en “machete” durante el 2016 fue de 0,4% teniendo su mayor pico en marzo (0,73%) (n= 362 estómagos). Aedo et al. (2007), menciona que en Chile el porcentaje de llenura en dicho periodo fue de 2,1% (n= 708 estómagos); estas diferencias pueden ocurrir debido a la diferencia en longitud de los peces, ya que en Chile se reportan ejemplares con longitudes mayores, además de las condiciones ambientales, zonas geográficas y tiempo de muestreo diferentes.
El número total de presas reportadas en este estudio fue de 39, correspondientes a los grupos zooplancton y fitoplancton. Estos valores son superiores a lo reportado en Chile, por Aedo et al. (2007) quienes indican que en el contenido estomacal de “machuelo” lo conforman 15 ítems presa pertenecientes a crustáceos, poliquetos, bivalvos, peces y fitoplancton. Estas diferencias indican una mayor demanda trófica por parte del machete de esta parte del Perú.
Los métodos de frecuencia de ocurrencia y numérico son usados para estimar la predominancia de presas de algún grupo en la dieta de los peces. Para E. maculatum, el grupo larvas es más importante en Chile (Aedo et al., 2007), el grupo Dinoflagelados y copépodos para la región La Libertad en Perú (Atoche, 2016), y el grupo Fitoplancton de manera general (Blaskovic et al., 2007; Cahuana, 1995; IMARPE, 2011). No obstante, estos métodos pueden subestimar la importancia de presas de gran tamaño y producir sesgos (Pillay, 1952; Windell, 1971; Hyslop, 1980; Konchina, 1991).
La estimación de biomasa húmeda de cada presa, a través del método volumétrico utilizado en este estudio es el primer paso hacia la estimación de contenido de carbono por presa. Si se estimaría el volumen por sí solo o a través del número de presas, tendríamos una sobre estimación de las diatomeas, como en el caso de la sardina Sardinops sagax, que pertenece a la familia Clupeidae igual que E. maculatum. Para este caso de estudio se sabe que la proporción diatomea: copépodo, 2:1 en volumen, es equivalente a 1:2.6 en carbono; por lo tanto, el zooplancton es una fuente de carbono superior al fitoplancton (Van der Lingen, 2002). Además, los copépodos tienen aproximadamente 8 veces más nitrógeno que las diatomeas, en comparación con volúmenes equivalentes entre ambos (Smayda, 1978, Hitchcock 1982).
La composición de la dieta por intervalos de tallas (21,0 - 24,0), (25,0 - 28,0) y (29,0 - 32,0) demostró que tanto los peces jóvenes como adultos se alimentan de las mismas presas; en contraste con Aedo et al. (2007) quienes encuentran que el “machete” tiene preferencia en cuanto a su alimentación dependiendo de la talla (< 28), (28 - 38). Llanos et al. (1996) mencionan que el contenido estomacal de “machetes” más jóvenes está compuesta por presas pequeñas tales como huevos de anchoveta, copépodos, nauplios, dinoflagelados y larvas de moluscos; en los adultos ingieren presas más grandes como los copépodos. Cabe mencionar que el comportamiento alimentario es característico de cada especie e incluso pueden variar aún entre individuos de la misma especie, de acuerdo a la localidad, la estación climática y la edad o el sexo (Prejs y Colomine, 1981).
Los resultados de este estudio muestran un aporte al conocimiento de dinámica alimentaria, alimento y hábitos alimentarios a través de tres métodos de estimación. Es necesario que se siga recopilando información para observar patrones de variación interanual. Además, se sugiere ampliar la escala de estudio, para observar variaciones latitudinales; y analizar otras especies con las que E. maculatum comparte hábitat, para conocer el hábitat trófico.
5. CONCLUSIONES
Ethmidium maculatum “machete” tiene mayor actividad alimentaria en el último semestre del año. Se alimenta de diatomeas, dinoflagelados, silicoflagelados y copépodos, sin embargo, las presas más abundantes en biomasa húmeda fueron del grupo copépodos. No presenta variación temporal en el consumo de plancton, pero sí de copépodos. No hay evidencia estadística para demostrar variación ontogénica en el consumo de presas.
6. AGRADECIMIENTOS
Al Ing. Luis Escudero Herrera, coordinador del Laboratorio Costero de Huanchaco en el periodo cuando se desarrolló el estudio, por brindar las facilidades para el uso de materiales y equipos utilizados. También, a la Blga. Betsy Buitrón Díaz, actual coordinadora del Laboratorio Costero de Huanchaco, por la gestión efectiva en la coordinación IMARPE-UNT, al finalizar la investigación. Esta investigación formó parte de la tesis: “Alimento y hábitos alimentarios Ethmidium maculatum “machete” en la Región La Libertad durante el año 2016” de la Blga. Pesq. Margot Ruíz García, desarrollada por el Laboratorio Costero de Huanchaco (IMARPE) y la escuela de Biología Pesquera de la Universidad Nacional de Trujillo.
7. CONTRIBUCIÓN DE LOS AUTORES
Dennis Atoche-Suclupe: Encargado de coordinar los muestreos, asesoramiento en el procedimiento de muestreo de contenido estomacal, tratamiento y análisis de datos del método volumétrico.
Zoila Culquichicón Malpica: Investigadora encargada de asesorar en la obtención de resultados utilizando los métodos frecuencia de ocurrencia y numérico
Margot Ruíz García: Apoyo en la realización de muestreos biológicos, reconocimiento de presas e índices de dinámica alimentaria.
8. CONFLICTO DE INTERESES
Los autores declaran que no existe conflicto de interés
9. FINANCIAMIENTO
Instituto del Mar del Perú – Oficina General de Promoción y Desarrollo de la Investigación de la Universidad Nacional de Trujillo (OGPRODEIN) – Recursos propios
10. REFERENCIAS BIBLIOGRÁFICAS
Aedo, G., Veas, R. Cubillos, L; Castro, L., Landaeta, M., Araya, M., Galleguillos, R., y Pedraza, M. (2007). Estudio biológico-pesquero de bacaladillo (Normanichthys crockeri) y machuelo (Ethmidium maculatum) en la zona centro-sur de chile. Universidad de concepción. Departamento de oceanografía. Informe final
Atoche, D. (2016). Seguimiento de la pesquería demersal y costera en la región La Libertad, 2016. Informe interno anual, IMARPE sede Huanchaco, 38 p.
Atoche, D. (2014). Seguimiento de la Pesquería Demersal Costera en la Región La Libertad, 2014. Informe interno anual, IMARPE sede Huanchaco, 37 p.
Atoche, D., y Culquichicón, Z., (2013). Biología y Pesquería de Ethmidium maculatum “machete” en la región La Libertad durante el 2010. SCIÉNDO, 16(2):27-42
Balech, E. (1988). Los Dinoflagelados Atlántico Sudoccidental. Publ. Espec. Inst.Esp.Oceanogr. 299 p.
Blaskovic, V., Castillo, D., Fernández, C., y Navarro, I. (2007). Hábitos alimentarios de las principales especies costeras del litoral de tumbes en el 2007. Informe técnico anual. Instituto del Mar del Perú. Tumbes – Perú, 13 p.
Boltovskoy, D. (1981). Atlas del zooplancton del Atlántico Sudoccidental y métodos de trabajo con el zooplancton marino. Publicación Especial del INIDEP, Mar de Plata, 936p.
Cahuana, A. (1995). Estudio de algunos aspectos biológicos del “machete” Ethmidium maculatum en las zonas de Callao e Ilo. Recuperado el 17 de octubre del 2020, de: http://www.imarpe.pe/imarpe/archivos/tesis/imarpe_restes_cahuana_quino_ruth_1995.pdf
Chirichigno, N. (1974). Clave para identificar los peces marinos del Perú. Inf. Inst. Mar Perú,44: 387 p.
Choy, C., Popp, B., Hannides, C., y Drazen, J. (2015). Trophic structure and food resource of epipelagic and mesopelagic fishes in the north Pacific Subtropical Gyre ecosystem inferred for nitrogen isotopic compositions. Limnology and oceanography, 60(4):1156 -1171.
Cupp, E. (1943). Marine Plankton Diatoms of the West Coast of North America. Bulletin of the Scripps Institution of Oceanography, 5(1): 237 p.
De Silva, S. (1973). Food and feeding habits of the herring Clupea harengus and the sprat C. sprattus. In: Inshore waters of the West coast of Scotland. Marine Biology, 20:282-290
Hyslop, E. (1980). Stomach contents analysis: a review of methods and their application. Journal of Fish Biology, 17:411–429.
Hitchcock, G. (1982). A comparative study of the size-dependent organic composition of marine diatoms and dinoflagellates. J. Plankt. Res, 4:363-377.
IMARPE. (2011). Anuario Científico Tecnológico IMARPE. Instituto del Mar del Perú (ISSN 1813 -2103). Callao - Perú.
IMARPE. (1996). Compendio biológico tecnológico de principales especies hidrobiológicas comerciales del Perú, 141p.
Khal’ko, V., Kiyashko, V., Khal’lko N. (2009). Circadian Rhythmic Changes in the Lipid Composition of Muscles in Kilka Clupeonella cultriventris (Clupeidae, Clupeiformes) during Natural Periodicity of Feeding in the Feeding and Spawning Periods in the Rybinsk Reservoir. Journal of Ichthyology, 49 (7):527–539.
Konchina, Y. (1991). Trophic status of the Peruvian anchovy and sardine. Journal of Ichthyology, 31:59–72.
Link, J., y Almeida, F. (2000). An overview and history of the food web dynamics program of the Northeast Fisheries Science Center, Woods Hole, Massachusetts. NOAA Technical Memorandum NMFS-NE 159, 1–60
Llanos, A., Herrera, G., y Bernal, P. (1996). Análisis del tamaño de las presas en la dieta de las larvas de cuatro clupeiformes en un área costera de Chile central. Sci. Mar. 60: 435-442p.
Nikolski, C. (1963). The ecology of fishes. Academic Press, London, 352 p.
Pillay, T. (1952). A critique of the methods of study of food of fishes. Journal of the Zoological Society of India, 4(1):185-200.
Pitt, T. (1973). Food of American plaice (Hippoglosooides platessoides) from the Grand Bank, Newfoundland. J. Fish. Res. Board. Can, 30(9):1261-1273.
Prejs, A., y Colomine, G. (1981). Métodos para el estudio de los alimentos y las relaciones tróficas de los peces. U. Central de Venezuela/U. de Varsovia. Caracas
Ramos, J. (1979). Fisiología de la reproducción y biología del lenguado Solea solea. Tesis doctoral de la Universidad Complutense de Madrid, 264 p.
Ruíz, M. (2016). Alimento y hábitos alimentarios Ethmidium maculatum “machete” en la Región La Libertad durante el año 2016. Tesis para optar el título de Biólogo Pesquero de la Universidad Nacional de Trujillo, 35 p
Santander, H. (1981). The zooplankton in an upwelling area off Peru. In: Richards, F.A. (Ed.), Coastal Upwelling Coastal and Estuarine Sciences. American Geophysical Union, Washington, DC. (1), 411–416p.
Shaw, A., Frazier, B., Kucklick, J., y Sancho, G. (2016). Trophic ecology of a predatory community in a shallow-water, high-salinity estuary assessed by stable isotope analysis. Marine and Coastal Fisheries, 8(1):46-61.
Smayda, T. (1978). From phytoplankters to biomass. In Phytoplankton Manual. Sournia, A. (Ed.). Unesco Monographs on Oceanographic Methodology, 6:273–279.
Sun, J. y Liu, D. (2003). Geometric models for calculating cell biovolume and surface area for phytoplankton. Journal of Plankton Research, 11 (25):1331–1346
Taylor, M. (2008). The Northern Humboldt Current Ecosystem and its resource dynamics: Insights from a trophic modeling and time series anaylisis. Ph.D. Thesis.Univ. Bremen.
Tresierra, A., Culquichicón, Z. (1995). Manual de Biología Pesquera. Editorial Libertad E.I.R.L. Trujillo. Perú, 226 p.
Van der Lingen, C. (2002). Diet of sardine Sardinops sagax in the southern Benguela upwelling ecosystem. South African Journal of marine Science 24:301- 316.
Whitehead, P. (1985). FAO species catalogue. Vol. 7. Catalogue critique des types de Poissons du Museum national d'Histoire naturelle. (Suite). Ordre des Clupeiformes (Familles des Clupeidae, Engraulididae et Denticipitidae). Bull. Mus. Natl. Hist. Nat. Ser. 4:1-77.
Windell, J. (1971). Food analysis and rate of digestion. In Methods for assessment of Fish Production in Fresh Waters (W.W. Ricker, ed). Oxford: Blackwell Scientific Publications, 215-226
Wootton, R. (1990). Ecology of teleost fishes. First Edition. Kluwer Academic Publishers. The Netherlands, 386 p.
Wootton, R. (1979). Energy cost of egg production and environmental determinants of fecundity in teleost fishes. Symp. Zool. Soc. Lond. 44:133-159.
Yuneva, T., Zabelinskii, S., Datsyk, N., Shchepkina, A., Nikolsky V., Shulman G. (2016). Influence of Food Quality on Lipids and Essential Fatty Acids in the Body of the Black Sea Sprat Sprattus sprattus phalericus (Clupeidae). Journal of Ichthyology. 56(3):397-405
Citar como:
Atoche-Suclupe, D.; Culquichicón, Z.; Ruíz, M. 2020. Aspectos tróficos de Ethmidium maculatum “machete”, región La Libertad, 2016. REBIOL 42(2):218-230. DOI: http://dx.doi.org/10.17268/rebiol.2020.40.02.09.